新冠病毒、中东病毒的共同秘密(小结篇)
接续:
新冠病毒、中东病毒的共同秘密(下)
由于之前的两篇文章都篇幅较长,信息量较大,因此,通过本文作一个简明的概括、总结。
SARS-CoV-2 2019年11、12月现身于武汉,MERS-CoV 2012年9月现身于沙特。两种病毒的现身地远隔重洋、相距万里,它们原本应该素昧平生、从未谋面、向无交集。
顺便有请诸位思考以下问题。SARS-CoV-2为什么偏偏出现在武汉?这是偶然的巧合,还是特意的选择?是某所泄漏了SARS-CoV-2,还是有人营造假象、栽赃嫁祸,转移视线、瞒天过海?如果是后者,为什么这么做?核心目的是什么?
这组问题不难想到,然而很少有人提起;“SARS-CoV-2偏偏出现在武汉”是一个重大疑点,上述问题可能是破解、查明SARS-CoV-2来源真相、疫情发生真相的关键所在和突破口,然而,耐人寻味的是,很多人对此讳莫如深、绝口不提,从不深究。
真相往往就隐藏在被刻意忽略、被刻意回避的细节之中。
是假象就会有破绽,作案者、制造假象者难免露出马脚。有没有发生过泄漏?疫情究竟是不是某所泄漏引起的?笔者作过两个分析,简明分析参见:
科学界为什么,为谁掩盖新冠病毒来源真相?(上)
一文“谎言一”小节的“两个疫情基本事实”部分;详细分析参见:
武汉病毒研究所是否泄漏了新冠病毒
一文。
言归正传。
SARS-CoV-2是β谱系B支系(乙型冠状病毒属Sarbe亚属)冠状病毒,MERS-CoV 是β谱系C支系(乙型冠状病毒属Merbe亚属)冠状病毒,虽然隶属同一谱系(属),但两者基因组差异非常巨大,进化距离非常遥远。SARS-CoV-2、MERS-CoV全基因组序列(核苷酸序列)仅28%可比对(Blast比对结果中的Query Cover值为28%),可比对部分相似度为67.06%;两种病毒Spike蛋白aa序列(amino acid sequence,氨基酸序列)76%可比对,可比对部分相似度仅为34.91%;在决定受体结合能力的Spike蛋白RBD(receptor binding domain,受体结合域)部分,两种病毒aa序列相似度同样非常低,仅为19.1%。
虽然发现地远隔重洋、相距万里,虽然素昧平生、向无交集,虽然基因组差异非常巨大,进化距离遥不可及,但SARS-CoV-2、MERS-CoV却如同心有灵犀的跨时空灵魂知己,如同无比默契的同门师兄弟,它们在感染、致病的关键环节、关键位点,存在诸多极低概率的,极为反常、离奇的,极度不可思议的结构、能力巧合。
DPP4结合能力、利用能力巧合
DPP4,dipeptidyl peptidase IV,二肽基肽酶4,是一种非常重要的人体丝氨酸蛋白酶兼多功能蛋白。
这组巧合包括以下4项具体巧合:
1、SARS-CoV-2、MERS-CoV都具有DPP4结合能力;
2、SARS-CoV-2、MERS-CoV都能以DPP4为受体感染人体细胞;
值得注意的是,SARS-CoV-2的“近亲”病毒,SARS-CoV-2同支系的众多冠状病毒,无一具有人类或动物DPP4结合能力,无一能以DPP4为受体感染人体或动物细胞(蛋白结合能力是以该蛋白为感染受体的先决条件)。
而且,以DPP4为感染受体,或具有DPP4结合能力的病毒非常罕见,截止2020年,具有DPP4结合能力的病毒可能只有4种:艾滋病病毒HIV-1、扁颅蝠冠状病毒HKU4、MERS-CoV、SARS-CoV-2 。因此,SARS-CoV-2、MERS-CoV 共同拥有的DPP4结合能力、以DPP4为感染受体的能力,是一种非常罕见的病毒能力。
HIV-1能以gp120蛋白(HIV-1的包膜蛋白—gp160蛋白的受体结合亚基)结合DPP4,但笔者未见有论文将DPP4称为HIV-1的感染受体;HKU4适宜结合蝙蝠DPP4,对人类DPP4结合能力微弱,它只能感染蝙蝠细胞,不能感染人体细胞。因此,在大千世界不计其数的病毒(包括冠状病毒和非冠状病毒)中,能以DPP4为受体感染人体细胞的病毒可能仅有两种,就是SARS-CoV-2和MERS-CoV。
还不止如此。
3、SARS-CoV-2、MERS-CoV在结合DPP4时锚挂的4个DPP4关键氨基酸位点相同。
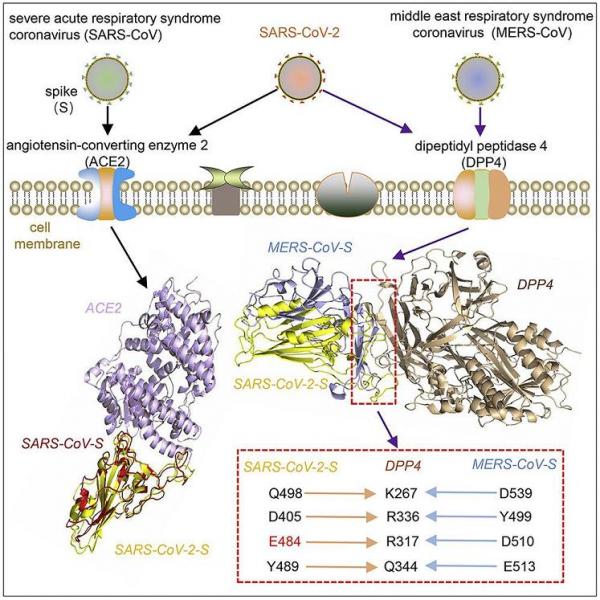
SARS-CoV 、SARS-CoV-2 、MERS-CoV 受体结合能力对照、关联图。
如图所示,SARS-CoV-2 Spike、MERS-CoV Spike 在结合DPP4时锚挂相同的4个DPP4关键氨基酸(K267、R336、R317、Q344)。
4、MERS-CoV在结合DPP4时共锚挂了14个DPP4氨基酸位点,巧合的是,DPP4在人体内的结合伴侣--人类腺苷脱氨酶(ADA)在结合DPP4时也锚挂了共14个DPP4氨基酸位点;令人震惊的是,而MERS-CoV、人类腺苷脱氨酶(ADA)所锚挂的14个DPP4氨基酸位点居然完全相同、一个不差!
MERS-CoV、人类腺苷脱氨酶(ADA)锚挂的14个共同的DPP4氨基酸位点,按照论文依据指出它们时的顺序依次是:1-R336、2-Q286、3-T288、4-R317、5-Q344、6-K267、7-L294、8-Y322、9-A291、10-L294 、11-I295、12-V341、13-I346、14-N229。
人类DPP4共有766个氨基酸位点。在766个氨基酸位点中恰好锚挂了14个相同的氨基酸位点,这实在是太过于巧合、太过于离奇、太过于匪夷所思了!
上述巧合表明:SARS-CoV-2、MERS-CoV极有可能都参照、借鉴、模仿了人类腺苷脱氨酶(ADA)的DPP4结合机制。MERS-CoV完全照搬、复用了ADA的14个DPP4结合/锚挂位点,SARS-CoV-2可能作了一些调整、变通,只复用(或至少复用)了ADA 14个DPP4结合/锚挂位点中的4个最关键位点。
S1/S2处氨基酸序列模式巧合
S1/S2表示冠状病毒Spike蛋白S1亚基、S2亚基的交界处。S1/S2位点是冠状病毒感染、致病的关键位点。
SARS-CoV-2、MERS-CoV S1/S2位点左侧的氨基酸序列模式高度相似。

4种病毒S1/S2(箭头指示处)位点附近的氨基酸序列
注:图中的SARS-CoV、SARSr-CoV RaTG13都是同谱系、同支系的SARS-CoV-2“近亲”病毒。
如上图所示,SARS-CoV-2、MERS-CoV S1/S2位点左侧氨基酸序列的高度相似性至少体现在以下3个方面:
1)、SARS-CoV-2 S1/S2位点左侧存在两种“近亲”所欠缺的独特四氨基酸插入PRRA,近亲欠缺,MERS-CoV 不缺,MERS-CoV S1/S2位点左侧恰巧存在与PRRA对应、匹配的四氨基酸PRSV。
2)、二者的四氨基酸组合PRRA、PRSV皆以PR(脯氨酸-精氨酸)开头,皆以非极性氨基酸结尾(A-丙氨酸、V-缬氨酸都是非极性氨基酸)。
3)、SARS-CoV-2、MERS-CoV在S1/S2处都具有furin酶切位点/furin切割位点(furin cleavage site,FCS)。PRRA、PRSV之后同为精氨酸R,RRA与R构成了SARS-CoV-2的furin切割序列RRAR,RSV与R则构成了MERS-CoV的furin切割序列RSVR。
对比鲜明且极为反常的是,SARS-CoV-2的所有“近亲”病毒,SARS-CoV-2同支系的所有冠状病毒,无一例外,在S1/S2处全都空缺与PRRA 对应的氨基酸,无一例外,在S1/S2处全都没有furin切割序列/furin酶切位点!如以下两图所示。
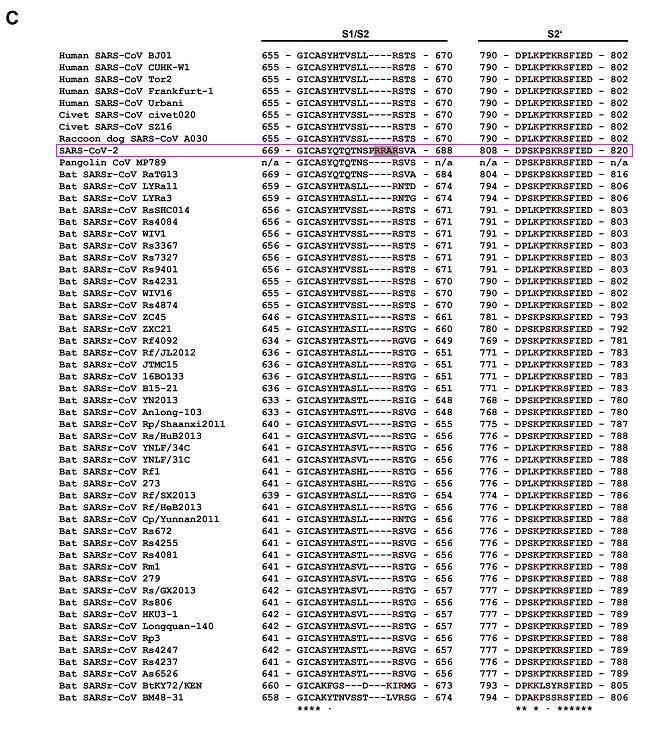
β谱系B支系部分人类、果子狸、貉、穿山甲、蝙蝠冠状病毒局部氨基酸序列比对
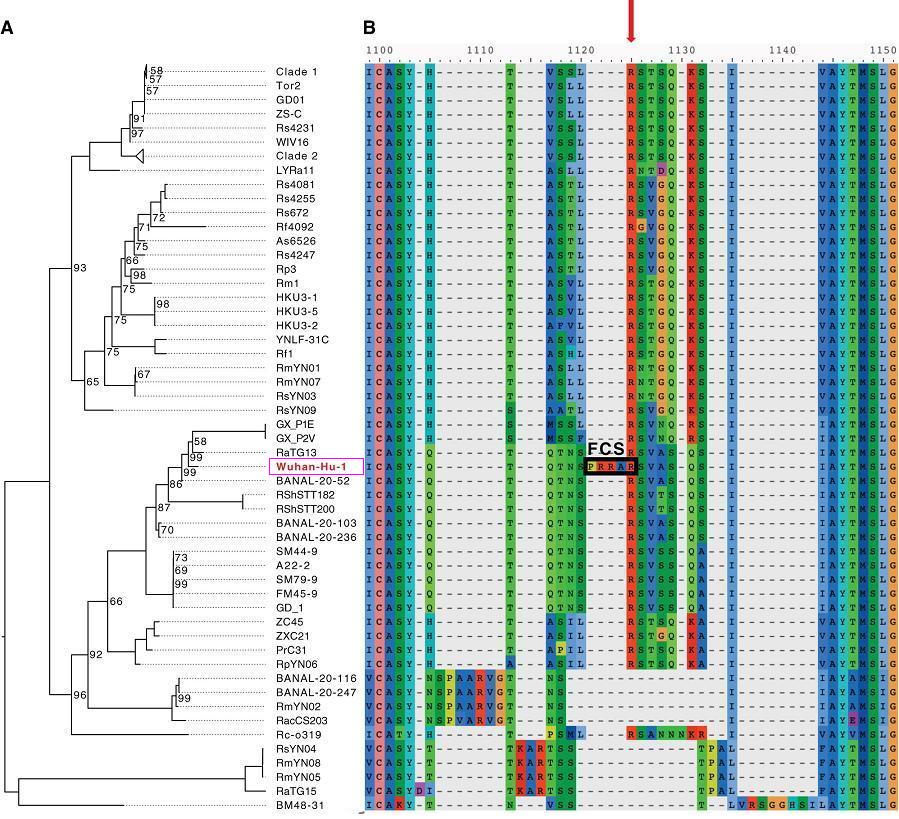
SARS-CoV-2(Wuhan-Hu-1毒株)与进化树中冠状病毒S1/S2位点附近的aa序列比对
由以上两图可见,SARS-CoV-2 S1/S2位点左侧独特的PRRA插入在图中极为醒目、突兀、异常,两图中的其它病毒全都空缺与PRRA对应的氨基酸。
SARS-CoV-2 S1/S2处氨基酸序列的高度反常还在于,Spike与SARS-CoV-2 spike同源性(相似度)大于40%的冠状病毒,无一具有furin 切割位点,当然更不具有S1/S2处的furin切割位点或furin切割序列。
P5位点脯氨酸巧合
SARS-CoV-2、MERS-CoV furin切割序列P5位点皆为脯氨基酸(P)。无比神奇、极度不可思之处在于,在大千世界不计其数的众多病毒(包括冠状病毒、非冠状病毒)中,具有furin切割位点,且furin切割序列P5位点为脯氨酸(P)的病毒仅有两种,这两种病毒恰巧就是SARS-CoV-2、MERS-CoV!
这是何等的默契,何等的心有灵犀!

SARS-CoV-2 、MERS-CoV furin切割序列P5位点皆为脯氨酸(P)
基本furin切割序列RXXR(如RRAR、RSVR)的四个氨基酸位点从右至左依次被称为P1、P2、P3、P4位点,P5位点是RXXR左侧最邻近的氨基酸位点。
P5位点使用脯氨酸是一个富有深意的设计,对病毒的感染、致病能力有多重作用(详情见上一篇文章对应小节),包括下面即将提到的,用于构成脯氨酸定向激酶作用基序和磷酸化位点。
P6、P5位点脯氨酸定向激酶作用基序巧合
脯氨酸定向激酶(proline-directed kinases)有两个二元(双氨基酸)作用基序:SP、TP。SARS-CoV-2、MERS-CoV furin切割序列P6、P5位点恰好分别使用了这两个作用基序,SARS-CoV-2使用了SP(丝氨酸-脯氨酸)基序,MERS-CoV则使用了另一个作用基序TP(苏氨酸-脯氨酸)。
这又是何等的默契,何等的心有灵犀!

MERS-CoV、SARS-CoV-2 P6、P5位点的脯氨酸定向激酶作用基序TP、SP
P6位点是P5位点左侧的下一个氨基酸位点。
由于furin切割序列P5位点为脯氨酸(P)的病毒仅有两种,因此,furin切割序列P6、P5位点为脯氨酸定向激酶作用基序(SP、TP)的病毒同样仅有两种,还是MERS-CoV、SARS-CoV-2。
MERS-CoV、SARS-CoV-2为什么要使用脯氨酸定向激酶作用基序?因为脯氨酸定向激酶作用基序构成病毒的磷酸化位点,基序中的丝氨酸或苏氨酸可被脯氨酸定向激酶磷酸化。病毒可通过磷酸化位点干预、改变人体正常的磷酸化机制,扰乱、破坏正常生理过程,导致种种疾病。病毒还可通过磷酸化有关蛋白质,干预、劫持人体信号通路、信号传导,阻滞/停滞细胞周期,使细胞处于有利于病毒复制、病毒生存的周期阶段。
蛋白质磷酸化异常,包括磷酸化途径的改变会导致多种严重疾病,尤其是癌症。SARS-CoV-2具有高度致癌性,SARS-CoV-2的致癌途径异常丰富。其它与磷酸化异常有关的疾病包括糖尿病、心血管疾病、阿尔茨海默症(Alzheimer's disease)、炎症性疾病等等。
上述种种极低概率、极度离奇、极度诡异、极度不可思议的巧合,不是随机性、无目的自然变异、自然演化所能解释、所能敷衍的,它们铁证如山地揭示、证明:SARS-CoV-2、MERS-CoV都是人为设计、制造的实验室病毒;两种病毒的设计思路高度一致、高度契合、一脉相承,它们极有可能出自同一实验室、同一团队或同一团伙之手。
(未完待续)